Психология педагогика : Контрольная работа: Физиология и питание растительных организмов
Контрольная работа: Физиология и питание растительных организмов
1. В чем
сущность автотрофного и гетеротрофного питания растительных организмов? Каковы
типы гетеротрофного питания?
По характеру
пищи, используемой в процессе жизнедеятельности, все живые организмы делятся на
автотрофных и гетеротрофных. Неорганические составные — CO2, H2O
и др. — служат основной пищей для автотрофных организмов (большинство
растений), которые синтезируют из них путём фотосинтеза или хемосинтеза органические
вещества: белки, жиры, углеводы, — составляющие пищу гетеротрофных организмов
(ряд растений, все Грибы, животные и человек). Помимо белков жиров и углеводов
гетеротрофным организмам необходимы витамины, нуклеиновые кислоты и микроэлементы.
Автотрофы
организмы, синтезирующие из неорганических соединений органические вещества.
Часть организмов (фотоавтотрофы) использует для этого энергию солнца. К ним
относятся высшие растения (исключение составляют растения-паразиты), водоросли
(фотоавтотрофные протисты), фотосинтезирующие бактерии. Они получают энергию в
ходе фотосинтеза, осуществляющегося в хлоропластах (эукариоты) или на клеточных
мембранах (прокариоты). В ходе фотосинтеза образуется не только глюкоза, но и аминокислоты,
используемые для построения белков. Другие организмы используют для этого
энергию, высвобождающуюся в ходе химических реакций. Такие организмы называются
хемоавтотрофами. К хемотрофам относятся хемосинтезирующие бактерии, образующие
органику в ходе хемосинтеза. Автотрофы являются продуцентами в сообществах,
именно они составлют первый ярус в пищевой пирамиде (первые звенья пищевых
цепей)
Гетеротрофы
организмы, которые не способны синтезировать органические вещества из
неорганических. Для синтеза необходимых для своей жизнедеятельности
органических веществ им требуются органические вещества, произведённые другими
организмами. В процессе пищеварения пищеварительные ферменты расщепляют
полимеры органических веществ на мономеры. В сообществах гетеротрофы — это
консументы различных порядков и редуценты.
Некоторые
организмы (например, хищные растения) сочетают в себе признаки как автотрофов,
так и гетеротрофов. Такие организмы называются миксотрофами.
У
низших организмов выделяют три типа питания продуктами в основном растительного
происхождения, т.е. гетеротрофного питания: симбиотический, паразитический и
сапрофитный.
При
симбиотическом питании один организм питается отходами другого, не причиняя ему
вреда. Например, нитрифицирующие бактерии, живущие на бобовых растениях
снабжают их азотом. В кишечнике млекопитающих находятся бактерии, помогающие
расщеплять питательные вещества, например кишечная палочка E.coli. Благодаря
безвредности данной бактерии для человека она широко используется при создании
БСС.
При
паразитическом питании организм-паразит разрушает системы жизнедеятельности
организма-хозяина.
При
сапрофитном питании организмы выделяют ферменты на мертвый или
разлагающийся органический материал. К ним относятся грибы, ряд бактерий и
насекомых. Некоторые сапрофиты выделяют ферменты протеазы, способные разлагать
белки, растворять оболочки других клеток, в том числе болезнетворных. Поэтому протеазы
широко применяют в качестве объектов биотехнологии в моющих средствах, а также
в БСС для обнаружения с помощью ферментативных реакций различных специфичных
для них белков-субстратов (Блукет и др., 2001).
2.
Перечислить меристемные ткани; указать их местоположение в теле растения и те
постоянные ткани, которые они образуют. Объяснить, почему одни из меристем
называются первичными, а другие – вторичными
Меристемы (греч.
meristos - делимый, делитель) стоят особняком среди других тканей, поскольку
состоят из живых недифференцированных клеток, способных постоянно делиться. В
онтогенезе этот тип растительных тканей возникает первым в результате деления
зиготы. На ранних этапах развития весь зародыш состоит из меристем. Затем из
них образуются все ткани растения.
Процесс
детерминации (лат. determinare - определять) происходит следующим образом.
Сначала в клетке возникает полярность. Она выражается в неодинаковой
организации полюсов клетки. Полярность наблюдается уже в яйцеклетке, которая
находится в зародышевом мешке. Так, у нее выделяют халазный полюс и полюс,
обращенный к пыльцевходу (микропиле). Полярность меристематических клеток,
которые детерминируются в клетки различных тканей, определяется различными
факторами окружающей среды. Такие факторы могут иметь физическую природу
(температура, свет, сила притяжения Земли и т.д.) или химическую (ионы,
фитогормоны и другие вещества, выделяемые микроокружением). Детерминация
приводит к внутриклеточным биохимическим перестройкам, в результате чего клетка
получает способность развиваться специализированно, т.е. морфологически
дифференцироваться в специализированную клетку ткани, определенной детерминацией.
Следующим
этапом будет рост клеток. Этот процесс проходит удивительно согласованно. При
этом не происходит смещения клеточных стенок относительно друг друга и
цитоплазматические связи между соседними клетками сохраняются. В результате
дифференцировки клетка приобретает окончательные размеры и форму, но утрачивает
способность размножаться. Вероятно, митостатическое воздействие оказывает
микроокружение, так как выделение неделящихся клеток приводит к превращению ее
в делящуюся клетку каллуса.
           Сами меристемы состоят из
клеток двух типов. Клетки первого типа называются инициальными (лат. initialis
- начальный). Они способны неограниченно долго размножаться (аналогичные клетки
животных называются стволовыми). При этом после деления одна дочерняя клетка
сохраняет свойства инициальной, а другая, продолжая делиться, детерминируется
как клетка определенной ткани и приступает к начальным этапам дифференцировки.
Именно они представляют собой клетки второго типа. Сами меристемы состоят из
клеток двух типов. Клетки первого типа называются инициальными (лат. initialis
- начальный). Они способны неограниченно долго размножаться (аналогичные клетки
животных называются стволовыми). При этом после деления одна дочерняя клетка
сохраняет свойства инициальной, а другая, продолжая делиться, детерминируется
как клетка определенной ткани и приступает к начальным этапам дифференцировки.
Именно они представляют собой клетки второго типа.
Инициальных
клеток обычно бывает немного, иногда всего одна. Расположенные на апексах
(верхушках осевых органов) они часто имеют форму многогранника (по граням здесь
и происходят деления клетки).
Морфологически
клетки меристем отличаются от других рядом признаков. Внешне они представляют
собой похожие друг на друга относительно мелкие клетки с тонкими стенками,
способными к растяжению. Ядро занимает центральное положение, вокруг ядра
располагается сравнительно небольшое количество цитоплазмы. Характерно сильное
развитие гранулярного эндоплазматического ретикулума, что свидетельствует об
усиленном синтезе белков в клетке. Имеется много митохондрий, зато вакуоли
мелкие, под световым микроскопом они не различимы.
Образовательные
ткани в теле растения располагаются в разных местах, в связи с чем их делят на
несколько групп.
Верхушечные, или апикальные, меристемы
располагаются на верхушках (апексах) осевых органов — стебля или корня. С
помощью этих меристем вегетативные органы растений осуществляют свой рост в
длину. Если такие меристемы выделить из соответствующих органов культурных
растений и вырастить на питательной среде, можно получить посадочный материал,
не зараженный вирусами.
Латеральные
меристемы также
характерны для осевых органов, где располагаются концентрически, в виде муфты.
Специализированные клетки здесь располагаются как внутри (ближе к сердцевине),
так и снаружи (ближе к поверхности тела). Первичные латеральные меристемы
образуются из апикальных и дают начало всем тканям растения. В дальнейшем их
развитие у разных форм растений происходит по-разному. У древесных они
сохраняются в течение всей жизни в виде камбия, обеспечивая вторичное
утолщение. У травянистых форм латеральные меристемы быстро исчезают, поэтому
вторичное утолщение у них не происходит.
Интеркалярные
(лат. intercalate
- внедрять), или вставочные, меристемы происходят от верхушечных. Они
представляют собой группы клеток, еще способных размножаться, но уже вставших
на путь дифференциации. Инициальных клеток среди них нет, зато много
специализированных.
Интеркалярные
меристемы можно обнаружить в основании молодых листьев. Они характерны для
злаков, где располагаются в нижних частях междоузлий, окруженных влагалищем
листа. Эта особенность позволяет подниматься полегшим после сильного ветра
злакам - изгиб побега образуется именно в указанных выше местах.
Раневые
меристемы обеспечивают
восстановление поврежденной части тела. Они образуются из расположенных рядом с
поврежденными участками специализированных живых клеток. Регенерация начинается
с дедифференциации - обратного развития от специализированных клеток к
меристематическим. Вступившие в этот процесс клетки вновь обретают способность
делиться. Оказавшись на поверхности, они превращаются в феллоген, который, в
свою очередь, образует пробку, покрывающую поверхность раны.
В другом
случае дедифференцированные клетки, делясь, образуют рыхлую паренхиматозную
ткань - каллус (лат. callus - толстая кожа, мозоль). При определенных условиях
из него могут формироваться органы растения. Иногда из клеток каллуса могут
развиваться зародыши, которые впоследствии развиваются в самостоятельный
организм (это легко наблюдать на отрезанном листе бегонии, где зародыши будут
развиваться из эпидермальных клеток в области перерезанных жилок).
Первичные меристемы обладают
меристематической активностью, т. е. способны к делению изначально. В ряде
случаев способность к активному делению может вновь возникнуть и у клеток, уже
почти утративших это свойство. Такие «вновь» возникшие меристемы называют вторичными.
Первичными
меристемами являются апикальная, прокамбий и интеркалярная, вторичными — камбий
и феллоген, перицикл и раневая меристема (Родионова и др., 1990).
3.
Охарактеризовать функции и строение корневых волосков
Различные
части корня выполняют неодинаковые функции и характеризуются определенными морфологическими
особенностями. Поступление почвенного раствора в корень происходит
преимущественно через зону всасывания, поэтому чем больше поверхность этого
участка корня, тем лучше он выполняет свою основную всасывающую функцию. Именно
в связи с этой функцией часть клеток кожицы вытянута в корневые волоски длиной
0,1—8 мм. Корневые волоски появляются в виде небольших сосочков — выростов
клеток эпиблемы. Рост волоска осуществляется у его верхушки. Оболочка корневого
волоска растягивается быстро. По прошествии определенного времени корневой
волосок отмирает. Продолжительность его жизни не превышает 10—20 дней.
Почти
всю клетку корневого волоска занимает вакуоль, окруженная тонким слоем
цитоплазмы. Ядро располагается в цитоплазме возле верхушки волоска. Корневые
волоски способны охватывать частички почвы, как будто срастаются с ними, что
облегчает поглощение из почвы воды и минеральных веществ. Поглощению
способствует также выделение корневыми волосками различных кислот (угольной,
яблочной, лимонной, щавелевой), которые растворяют частички почвы.
Формируются
корневые волоски очень быстро (у молодых сеянцев яблони за 30—40 ч). На 1 кв.
мм корня при благоприятных условиях образуется до 300—400 корневых волосков,
которые создают огромную поглощающую поверхность. У одной особи
четырехмесячного растения ржи примерно 14 млрд. корневых волосков с площадью
поглощения около 400 м2 и суммарной длиной более 10 тыс. км;
поверхность всей корневой системы, включая корневые волоски, составляет
примерно 640 м2, т.е. в 130 раз больше, чем у побега. Функционируют
корневые волоски недолго — обычно 10—20 дней. Средняя медленность жизни волоска
у винограда — от 10 до 40 суток. Сменяют отмершие корневые волоски в более
нижней части корня новые. Таким образом, наиболее деятельная, всасывающая зона
корней все время перемещается вглубь и в стороны вслед за растущими кончиками
разветвлений корневой системы. При этом общая всасывающая поверхность корней
все время увеличивается.
Каждый
волосок представляет из себя вытянутую клеточку. В связи с ростом корня,
перемещением и обновлением поглощающей зоны происходит константа смена
волосков, обеспечивающая непрерывную активную работу корневой системы
(Андросов, 2006).
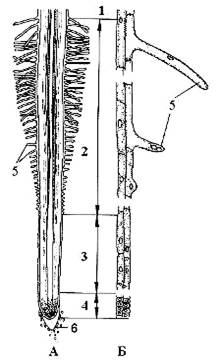
Рис. 1. Строение корня
проростка пшеницы: А - схема строения корня; Б - дифференциация клеток
ризодермы и экзодермы. 1 - зона проведения, 2 - зона всасывания, 3 - зона
растяжения, 4 - зона деления, 5 - корневой волосок, 6 - корневой чехлик
(Андросов, 2006).
4.
В чем отличие анатомического строения однолетней и многолетней ветки одного и
того же растения?
Анатомическое
строение типичного стебля соответствует его главным функциям. В нем развита
система проводящих тканей, часто очень сложная, которая связывает воедино все
органы растения; с помощью механических тканей стебель поддерживает все
надземные органы и выносит листья в благоприятные условия освещения.
Стебель, как
и весь побег в целом, представляет собой «открытую» систему роста, т.е. он
длительное время нарастает и на нем возникают новые органы. В стебле имеется
система меристем, поддерживающих нарастание тканей в длину и толщину. Стебель
существенно отличается от корня в том отношении, что формирование его тканей из
верхушечных меристем не имеет той строго акропетальной последовательности,
которая характерна для корня. Нарушение строго акропетального формирования
тканей в стебле объясняется тем, что на апексе побега с правильной
периодичностью возникают листовые примордии, что ведет к раннему вычленению
узлов, а развитие междоузлий запаздывает.
Часто рост
междоузлий и развитие в них постоянных тканей продолжаются длительное время за
счет работы тех остаточных интеркалярных меристем, которые сохраняются у
оснований нескольких междоузлий. Ярким примером такого интеркалярного
(вставочного) роста может служить стебель злака, у которого апикальная
меристема очень рано расходуется на образование соцветия, а быстрое вытягивание
(колошение) обязано именно вставочному росту. Таким образом, для стеблей
характерна сложная система меристем— верхушечных, боковых и вставочных.
В инициальном
кольце под молодыми листовыми зачатками возникает прокамбий. Внутрь от
прокамбия обозначается сердцевина, а наружу от него — первичная кора; самый
поверхностный слой верхушечной меристемы образует протодерму (и в дальнейшем
эпидерму).
В результате
деятельности первичных меристем складывается первичная структура стебля. Если
камбий работает слабо или совсем отсутствует, то она может сохраняться
длительное время. Если камбий энергично откладывает вторичные проводящие ткани,
то первичное строение быстро сменяется вторичным. Границу между ними иногда
провести трудно, так как клетки камбия начинают делиться еще до окончательного
формирования первичных тканей.
Камбиальный
слой откладывает внутрь вторичную ксилему, а сам растягивается на ее
поверхности и отодвигается от центра, оставаясь между ксилемой (древесиной) и
корой. Одновременно камбий откладывает наружу слои вторичной флоэмы (луба).
Первичная кора при длительном вторичном утолщении стебля погибает и заменяется
вторичной корой, под которой понимают совокупность всех вторичных тканей,
расположенных наружу от камбия. На поверхности стебля перидерма сменяет эпидерму.
В результате всех этих преобразований складывается вторичное строение стебля (Билич,
Крыжановский, 2005).
 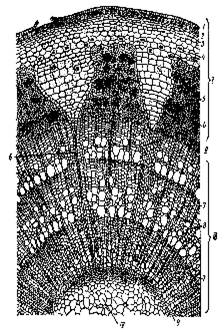
Рис. 2.
Поперечный разрез молодого стебля липы: К-камбий; Колл-колленхима; Мкс-
метаксилема; Мф-метафлоэма; П-паренхима; ПК- первичная кора; ПКс-первичная ксилема;
Пркс- протоксилема; Прф- протофлоэма; ПФ-первичная флоэма; С- сердцевина; Эп-
эпидерма. (Крамер, Козловский, 1983)
Рис. 3.
Разрез трехлетней ветки липы: 1-кора, II-камбий, III-древесина, IV-сердцевина; 1-
отмерший эпидермис, 2-перидерма, 3-колленхима, 4-клетки основной ткани с
друзами, 5-участки флоэмы, 6-сердцевинный луч, 7-осенняя древесина, 8- весенняя
древесина, 9-первичная ксилема. (Крамер, Козловский, 1983)
Развивающийся
стебель непременно в той или иной мере подвергается утолщению. В зависимости от
участия в этом процессе камбия утолщение может быть первичным или вторичным.
Первичное
утолщение представляет собой увеличение объема стебля за счет растяжения
клеток, составляющих первичные ткани. Этот процесс начинается уже в
непосредственной близости от конуса нарастания и может осуществляться
по-разному. Если преимущественно утолщается первичная кора, то утолщение
называется кортикальным (лат. cortex - кора), что можно наблюдать у кактусов.
Утолщение, которое осуществляется за счет сердцевины, называют медуллярным
(лат. medullaris, medulla — костный мозг, мозговой), оно имеет место, в
частности, при формировании картофельного клубня.
Вторичное
утолщение приводит к увеличению диаметра стебля, что неизбежно ведет к разрыву
и отмиранию первичной коры, на смену которой приходит кора вторичная, имеющая,
совершенно иное строение. Она состоит из образовавшейся вместо эпидермы
перидермы, остатков первичной коры и первичной флоэмы, а также вторичной
флоэмы. Впоследствии в результате деятельности камбия, регулярно откладывающего
вторичную флоэму, и феллогена, откладывающего пробку, формируется корка. У
некоторых растений, например у пробкового дуба, образуется очень мощный слой
корки, и человек широко использует ее в своей деятельности.
Вызванное
деятельностью камбия вторичное утолщение продолжается в течение всей жизни растения
и за много лет может привести к формированию ствола, имеющего колоссальный
объем и диаметр, превышающий 10 м. У деревьев, растущих в дождевых тропических
лесах, интенсивность деления камбиальных клеток очень мало изменяется в течение
года, поэтому у них почти не выражена слоистость древесины. Напротив, у
деревьев, произрастающих в условиях переменного климата, где сезонные колебания
температуры и влажности заметны очень хорошо, уровень деятельности камбия
находится в зависимости от времени года. Наибольшая интенсивность деления
инициальных клеток здесь отмечается весной, когда из почек интенсивно
развиваются молодые побеги, на которых имеются листья. В этот период в основном
откладываются проводящие воду элементы — трахеиды (большая часть древесины образована
именно ими) и членики сосудов, которые имеют большие внутренние объемы и тонкие
оболочки. Летом инициальные клетки камбия делятся реже и дают начало
толстостенным трахеидам и волокнам. У этих элементов функция проведения воды
выражена крайне слабо или отсутствует совсем, зато они прекрасно выполняют
функцию опоры. Осенью деятельность камбия ослабляется еще сильнее, а позднее
прекращается вовсе, зимой камбий не функционирует. В результате образовавшаяся
за год древесина неоднородна и ее можно подразделить на раннюю и позднюю, которые,
чередуясь, совместно формируют ежегодный слой прироста в виде кольца.
Ширина и
выраженность годичных слоев широко варьирует не только у представителей разных
видов растений, она неодинакова даже у одного и того же экземпляра и напрямую
зависит от влияния различных факторов окружающей среды. В годы с наиболее
благоприятными погодными условиями (следует отметить, что этот термин для
разных растений имеет совсем неодинаковый смысл - благоприятные погодные
условия для одной экологической группы могут быть весьма неприятными для
другой) слои прироста имеют наибольшую ширину, напротив, в неблагоприятные годы
образуются узкие слои. Таким образом, пересчитав все годичные кольца, можно
определить возраст дерева, измерив их толщину, иметь достаточно верное
представление о метеорологических условиях давно прошедших лет, учитывая, что
возраст наиболее старых деревьев может составлять не одно тысячелетие. Однако
здесь необходимо соблюдать некоторую осторожность, потому что в особо неблагоприятных
условиях годичные слои могут и не возникать. И наоборот, если летом у деревьев
вредителями уничтожались листья, после некоторого покоя камбий откладывает слой
древесины, по строению сходный с ранней весенней, что в итоге создает иллюзию
двух годичных слоев.
Теперь
становится понятным, что основная часть клеток многолетнего растения,
способного к вторичному утолщению, состоит из мертвых клеток, которые сообща
обеспечивают телу растения надежную опору (Матвеев, Тарабрин, 1989).
5.
Дать определение почки и описать типы почек по их положению и функциональному
значению
Почка
представляет собой орган побега, который обеспечивает его верхушечное
нарастание и ветвление. В состоянии покоя почка является зачаточным побегом, в
нем имеется сильно укороченная ось — зачаток стебля, заканчивающийся конусом
нарастания. На оси вегетативной почки располагаются зачаточные листья, причем в
их пазухах уже находятся зачатки пазушных почек следующего поколения. Листовые
примордии, определяющие нахождение узла на зачаточном стебле, располагаются
настолько близко друг к другу, что определить здесь междоузлия становится
довольно трудно.
Обычно
зимующие почки снаружи покрыты специализированными чешуями, представляющими
собой видоизмененные наружные листья. Они предохраняют почку от высыхания и
отчасти от перепадов температуры. Такие почки называются закрытыми. Если почка
лишена защитных чешуи, она называется открытой. Эти почки на самом деле не
лежат абсолютно «голо», а обычно бывают прикрыты прилистниками или листовыми влагалищами.
Часто одно и то же растение (как правило, многолетнее) летом имеет открытые
почки, а зимует с закрытыми. Многие тропические деревья, имеют только открытые
почки, а некоторые многолетние травы могут даже с такими почками зимовать.
По
расположению почки могут быть верхушечными и пазушными (рис. 4). Верхушечные
почки обеспечивают апикальное нарастание побега. Самую первую верхушечную почку
можно обнаружить еще в зародыше. Из нее в последующем будут
образовываться все органы побега. Пазушные почки находятся в пазухах листьев,
положение их на стебле напрямую зависит от листорасположения. Они формируются
экзогенно из апикальных меристем и обычно обнаруживаются в пазухе третьего
пятого листового примордия. В пазухе одного листа может находиться одна или
несколько почек В последнем случае почки располагаются либо рядом друг
с другом вокруг стебля (как, например, дочерние луковички в материнской
луковице чеснока), либо вертикально одна над другой (в частности, так
происходит у ежевики). Первый тип расположения называется коллатеральным, второй
- сериальным. Еще одним типом почек являются придаточные почки. В отличие от
пазушных, они закладываются эндогенно и развиваются не из апикальных меристем,
а из камбия, живых клеток паренхимы или эпидермы путем их дифференциации. Общий
план строения таких почек не отличается от других, но месторасположение может
быть самым разнообразным. Их можно обнаружить на стеблях, но, в отличие от пазушных
почек, придаточные никак не связаны с листьями и поэтому обычные почки образуются
на листьях. Такими почками размножается комнатное растение бриофиллюм
(каланхое). У бегоний придаточные почки развиваются из эпидермальных клеток,
расположенных поблизости от перерезанных жилок листа. Этот тип почек на листьях
могут также образовывать многие папоротники.
Кроме побега,
придаточные почки развиваются на корне. Развившиеся из них в дальнейшем побеги
называются корневыми отпрысками, которые для многих растений играют важную роль
в вегетативном размножении.
Многие
многолетние растения способны образовывать спящие почки. Они возникают как
пазушные или придаточные почки, но не развиваются в побег, а впадают в
состояние покоя и пребывают в нем в течение довольно длительного времени,
сохраняя при этом жизнеспособность. На самом деле покой спящих почек не
абсолютен, иначе образовавшиеся в результате вторичного роста ткани просто
закрыли бы их. Действительно, спящие почки растут, только очень медленно, образуя
за год несколько метамеров, при этом ось почки несколько утолщается и
увеличивает свою длину в соответствии с шириной образовавшегося годичного слоя.
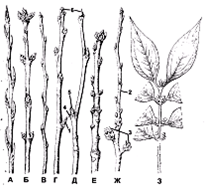 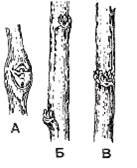 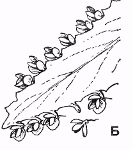
Рис. 4. Расположение
почек: А - тополь канадский (Populus canadensis); Б - сирень обыкновенная
(Syringa uulgaris); В - хурма кавказская (Diospyros lotus); Г - орех грецкий
(Juglans regia); Д - платан восточный (Piatanus orientalis); E - фисташканастоящая
(Pistacia vera); Ж - вишня обыкновенная (Cerasus uulgaris);3 - зимоцвет ранний
(Meratia praecox); 1 - цветочная почка; 2 - ростовой'побег; 3 - плодущие побеги
(плодушки); 4 - листовой след; 5 – пазушная почка; 6 - верхушечная почка
(Билич, Крыжановский, 2005).
Рис. 5. Расположение
почек: А-пазушное, коллатеральное; Б,В-пазушное снриальное (Билич, Крыжановский,
2005)
Рис. 6. Придаточные почки
на листьях у бриофиллюма (Билич, Крыжановский, 2005)
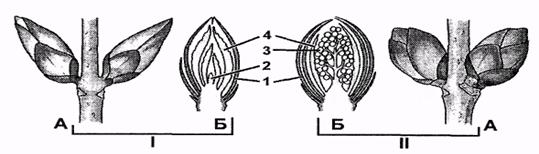
Рис. 7. Строение почек бузины: I
- вегетативные почки; II - генеративные почки; А - внешний вид и расположение
на стебле; Б — продольный разрез; 1 — почечные чешуи; 2 - конус нарастания; 3 - зачаточное соцветие;
4 – листья (Билич, Крыжановский, 2005)
Зачаточные
листья спящей почки трансформируются в чешуи, а образовавшиеся в них пазушные
почки позволяют ей ветвиться. Сигналом для выхода из состояния относительного
покоя спящей почки служит повреждение или снижение активности нормальных почек.
Хорошо известно появление молодой поросли на пнях или на стволах после сильной
обрезки ветвей. Спящие почки в основном характерны для цветковых растений,
причем встречаются не только у деревьев и кустарников, но и у многолетних трав.
Особенно они важны для кустарников, поскольку способствуют кущению. У голосеменных
спящие почки образуются значительно менее интенсивно, поэтому появление молодой
поросли на пнях ели или сосны обычно не происходит.
По
выполняемым жизненным функциям почки подразделяются на вегетативные, состоящие
из зачаточного стебля (оси) и зачатков листьев и генеративные, несущие также
зачатки соцветий и цветков. Из вегетативной почки образуется стебель с листьями
и почками, а из генеративной — одиночный цветок или соцветие (Васильев и др.,
1998).
6.
Доказать на конкретных примерах нетождественность биологических понятий «усы» и
«усики». Объяснить биологическое значение тех и других
"Усы"
- длинные наземные побеги, несущие дочерние розетки (например усы земляники,
живучки ползучей). Как полноценные стебли, усы способны образовывать на месте
соприкосновения с почвой корневую систему и надземную часть растения.
А
вот когда говорят, что у растения есть «усики», имеются ввиду длинные и тонкие
стебельки без всяких чешуек и листиков, на которых никаких розеток не
образуется. Усики у растений из семейства бобовых - у гороха, чины -
представляют собой окончание листа, его главный стержень с двумя - тремя
боковыми завивающимися спирально усиками. Когда в эту спираль попадается
какой-нибудь посторонний предмет, усик цепляется за него. Так, хватаясь усиками
листьев за соседние предметы, растущий стебель гороха и будет подниматься вверх
а без этого он не смог бы тянуться к свету и рано или поздно лег бы на землю.
Впрочем, верхушка стебля все равно будет стараться расти вверх, и в конце
концов усики где-нибудь нащупают опору и поднимут растение вверх. Усики у
небобовых растений чаще всего представляют собой стебли. Они никак не связаны с
листьями, разве только тем, что могут отходить от стебля в одном и том же
месте: где лист там и усик (немного выше черешка листа). Это хорошо видно у
растений семейства тыквенных, например у огурцов. Усики у них простые и не
имеют никаких ответвлений. У тыквы усики похожи на руку с тремя пальцами, а у
мексиканского огурца - и со всеми пятью пальцами. Интересно наблюдать за ростом
таких усиков: как только начинает разворачиваться очередной лист на растущем
конце стебля, так вместе с ним начинают расти стерженек усика и его
растопыренные "пальцы". Эти пальцы стараются охватить все, что
попадается на пути, и удлиняются, совершая своими кончиками круговые,
вращательные движения. Достигнув опоры, кончик усика закручивается вокруг нее.
Зацепившись, он начинает подтягивать к себе главный стебель и старается
свернуться спиралью. На усиках огурцов этот процесс видно очень хорошо.
На
этом активная деятельность усика прекращается. Свернутый спиралью он так и
живет, пока жив стебель (у трав) или пока не покроется твердой корой (у
древесных лиан). Усики, не нашедшие опоры, через 3-5 дней бессильно повисают,
отмирают и опадают. Чем больше разветвлений у усика, тем больше вероятность,
что хоть одно из них найдет себе зацепку. В этом отношении самые развитые,
самые совершенные усики у винограда. Но интересно то, что виноградные усики - это
не что иное как недоразвитое соцветие (Хржановский и др., 1988).
Библиография
1. Андросов П.В.
Ботаника. Учебник для агрономических факультетов сельскохозяйственных вузов. В
2 т. Т. 1 – Анатомия и морфология растений. Иркутск: изд-во «СИИБ», 2006. С.
112-142.
2. Билич Г.Л.,
Крыжановский, В.А. Биология. Полный курс. В 3 т. Т. 2. Ботаника. М.: ООО
«Издательство Оникс», 2005. С. 136-157.
2. Блукет Н.А., Емцев
В.Т., Воронович Е.В., Карпатова М.А. Ботаника с основами физиологии растений и
микробиологии. Учебник для высших учебных заведений с/х профиля. М.: Академия,
2001. С. 21-29.
3. Васильев А.Е., Воронин
Н.С., Еленевский А.Г. и др. Ботаника. Морфология и анатомия растений. М.:
Просвещение, 1998. С. 34-50.
4. Крамер П.В.,
Козловский Т.А. Физиология древесных растений. М.: Лесная промышленность, 1983.
С. 64-71.
5. Матвеева Г.В.,
Тарабрин А.Д. Ботаника. М.: Агропромиздат, 1989. С. 112-123.
6. Родионова А.С.,
Барчукова М.В. Ботаника. М.: Агропромиздат, 1990. С. 98-114.
7. Хржановский В.Г.,
Пономаренко С.Ф. Ботаника. М.: Агропромиздат, 1988. С. 14-38.
|



